Learninsta presents the core concepts of Biology with high-quality research papers and topical review articles.
Pre-Fertilization: Structures and Events
The hormonal and structural changes in plant lead to the differentiation and development of floral primordium. The structures and events involved in prefertilization are given below.
Male Reproductive part Androecium
Androecium is made up of stamens. Each stamen possesses an anther and a fiament. Anther bears pollen grains which represent the male gametophyte. In this chapter we shall discuss the structure and development of anther in detail.
Development of anther:
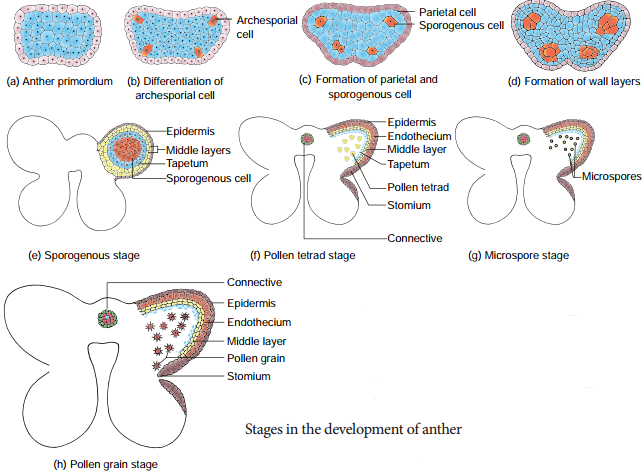
A very young anther develops as a homogenous mass of cells surrounded by an epidermis. During its development, the anther assumes a fourlobed structure. In each lobe, a row or a few rows of hypodermal cells becomes enlarged with conspicuous nuclei. This functions as archesporium.
The archesporial cells divide by periclinal divisions to form primary parietal cells towards the epidermis and primary sporogenous cells towards the inner side of the anther. The primary parietal cells undergo a series of periclinal and anticlinal division and form 2-5 layers of anther walls composed of endothecium, middle layers and tapetum, from periphery to centre.
Microsporogenesis:
Th stages involved in the formation of haploid microspores from diploid microspore mother cell through meiosis is called Microsporogenesis. The primary sporogeneous cells directly, or may undergo a few mitotic divisions to form sporogenous tissue. The last generation of sporogenous tissue functions as microspore mother cells. Each microspore mother cell divides meiotically to form a tetrad of four haploid microspores
(microspore tetrad).
Microspores soon separate from one another and remain free in the anther locule and develop into pollen grains. The stages in the development of microsporangia is given in Figure 1.4. In some plants, all the microspores in a microsporangium remain held together called pollinium.
Example: Calotropis. Pollinia are attached to a clamp or clip like sticky structure called corpusculum. The fiamentous or thread like part arising from each pollinium is called retinaculum. The whole structure looks like inverted letter ‘Y’ and is called translator.
T.S. of Mature anther
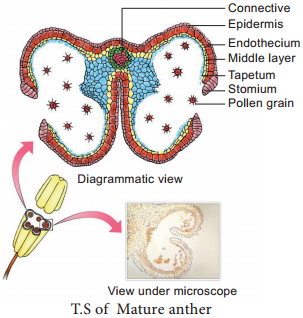
Transverse section of mature anther reveals the presence of anther cavity surrounded by an anther wall. It is bilobed, each lobe having 2 theca (dithecous). A typical anther is tetrasporangiate. The T.S. of Mature anther is given in Figure 1.5.
1. Anther wall
The mature anther wall consists of the following layers
a. Epidermis
b. Endothecium
c. Middle layers
d. Tapetum.
a. Epidermis:
It is single layered and protective in function. The cells undergo repeated anticlinal divisions to cope up with the rapidly enlarging internal tissues.
b.Endothecium:
It is generally a single layer of radially elongated cells found below the epidermis. The inner tangential wall develops bands (sometimes radial walls also) of α cellulose (sometimes also slightly lignified). The cells are hygroscopic. In the anthers of aquatic plants, saprophytes, cleistogamous flowers and extreme parasites endothecial diffrentiation is absent.
The cells along the junction of the two sporangia of an anther lobe lack these thickenings. Ths region is called stomium. Ths region along with the hygroscopic nature of endothecium helps in the dehiscence of anther at maturity.
c. Middle layers:
Two to three layers of cells next to endothecium constitute middle layers. They are generally ephemeral. They disintegrate or get crushed during maturity.
d. Tapetum:
It is the innermost layer of anther wall and attains its maximum development at the tetrad stage of microsporogenesis. It is derived partly from the peripheral wall layer and partly from the connective tissue of the anther lining the anther locule. Thus, the tapetum is dual in origin.
It nourishes the developing sporogenous tissue, microspore mother cells and microspores. The cells of the tapetum may remain uninucleate or may contain more than one nucleus or the nucleus may become polyploid.
It also contributes to the wall materials, sporopollenin, pollenkitt, tryphine and number of proteins that control incompatibility reaction .Tapetum also controls the fertility or sterility of the microspores or pollen grains. There are two types of tapetum based on its behaviour. They are:
Secretory tapetum (parietal/glandular/cellular):
The tapetum retains the original position and cellular integrity and nourishes the developing microspores.
Invasive tapetum (periplasmodial):
The cells loose their inner tangential and radial walls and the protoplast of all tapetal cells coalesces to form a periplasmodium.
Functions of Tapetum:
- It supplies nutrition to the developing microspores.
- It contributes sporopollenin through ubisch bodies thus plays an important role in pollen wall formation.
- The pollenkitt material is contributed by tapetal cells and is later transferred to the pollen surface.
- Exine proteins responsible for ‘rejection reaction’ of the stigma are present in the cavities of the exine. Thse proteins are derived from tapetal cells.
2. Anther Cavity:
The anther cavity is filed with microspores in young stages or with pollen grains at maturity. Th meiotic division of microspore mother cells gives rise to microspores which are haploid in nature.
3. Connective:
It is the column of sterile tissue surrounded by the anther lobe. It possesses vascular tissues. It also contributes to the inner tapetum.
Microspores and pollen grains
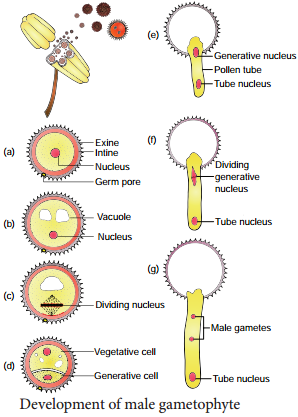
Microspores are the immediate product of meiosis of the microspore mother cell whereas the pollen grain is derived from the microspore. The microspores have protoplast surrounded by a wall which is yet to be fully developed. The pollen protoplast consists of dense cytoplasm with a centrally located nucleus. The wall is diffrentiated into two layers, namely, inner layer called intine and outer layer called exine.
Intine is thin, uniform and is made up of pectin, hemicellulose, cellulose and callose together with proteins. Exine is thick and is made up of cellulose, sporopollenin and pollenkitt. The exine is not uniform and is thin at certain areas.
When these thin areas are small and round it is called germ pores or when elongated it is called furrows. It is associated with germination of pollen grains. The sporopollenin is generally absent in germ pores. The surface of the exine is either smooth or sculptured in various patterns (rod like, grooved, warty, punctuate
etc.) Th sculpturing pattern is used in the plant identifiation and classifiation.
Shape of a pollen grain varies from species to species. It may be globose, ellipsoid, fusiform, lobed, angular or crescent shaped. The size of the pollen varies from 10 micrometers in Myosotis to 200 micrometers in members of the family Cucurbitaceae and Nyctaginaceae.
Pollenkitt is contributed by the tapetum and coloured yellow or orange and is chiefly made of carotenoids or flvonoids. It is an oily layer forming a thick viscous coating over pollen surface. It attracts insects and protects damage from UV radiation.
Development of Male gametophyte:
The microspore is the first cell of the male gametophyte and is haploid. The development of male gametophyte takes place while they are still in the microsporangium. The nucleus of the microspore divides mitotically to form a vegetative and a generative nucleus.
A wall is laid around the generative nucleus resulting in the formation of two unequal cells, a large irregular nucleus bearing with abundant food reserve called vegetative cell and a small generative cell.
Generally at this 2 celled stage, the pollens are liberated from the anther. In some plants the generative cell again undergoes a division to form two male gametes. The pollen is liberated at 2 celled stage. In 60% of the angiosperms pollen is liberated in 2 celled stage.
Further, the growth of the male gametophyte occurs only if the pollen reaches the right stigma. The pollen on reaching the stigma absorbs moisture and swells.
The intine grows as pollen tube through the germ pore. In case the pollen is liberated at 2 celled stage the generative cell divides in the pollen into 2 male cells (sperms) aftr reaching the stigma or in the pollen tube before reaching the embryo sac. The stages in the development of male gametophyte is given in Figure 1.6.
Female reproductive part Gynoecium
The gynoecium represents the female reproductive part of the flower. The word gynoecium represents one or more pistils of a flower. The word pistil refers to the ovary, style and stigma. A pistil is derived from a carpel. The word ovary represents the part that contains the ovules. The stigma serves as a landing platform for pollen grains. The style is an elongated slender part beneath the stigma. The basal swollen part of the pistil is the ovary. The ovules are present inside the ovary cavity (locule) on the placenta.
Gynoecium (carpel) arises as a small papillate outgrowth of meristematic tissue from the growing tip of the flral primordium. It grows actively and soon gets diffrentiated into ovary, style and stigma. The ovules or megasporangia arise from the placenta. The number of ovules in an ovary may be one (paddy, wheat and mango) or many (papaya, water melon and orchids).
Structure of ovule (Megasporangium):
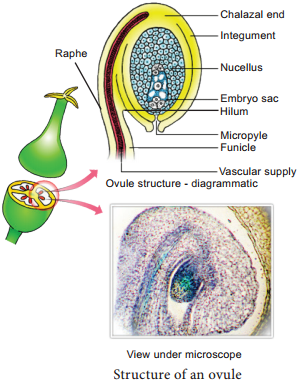
Ovule is also called megasporangium and is protected by one or two covering called integuments. A mature ovule consists of a stalk and a body. The stalk or the funiculus (also called funicle) is present at the base and it attaches the ovule to the placenta.
The point of attachment of funicle to the body of the ovule is known as hilum. It represents the junction between ovule and funicle. In an inverted ovule, the funicle is adnate to the body of the ovule forming a ridge called raphe. The body of the ovule is made up of a central mass of parenchymatous tissue called nucellus which has large reserve food materials.
The nucellus is enveloped by one or two protective coverings called integuments. Integument encloses the nucellus completely except at the top where it is free and forms a pore called micropyle. The ovule with one or two integuments are said to be unitegmic or bitegmic ovules respectively.
The basal region of the body of the ovule where the nucellus, the integument and the funicle meet or merge is called as chalaza.
There is a large, oval, sac-like structure in the nucellus toward the micropylar end called embryo sac or female gametophyte. It develops from the functional megaspore formed within the nucellus. In some species (unitegmic tenuinucellate) the inner layer of the integument may become specialized to perform the nutritive function for the embryo sac and is called as endothelium or integumentary tapetum (Example: Asteraceae).
There are two types of ovule based on the position of the sporogenous cell. If the sporogenous cell is hypodermal with a single layer of nucellar tissue around it is called tenuinucellate type. Normally tenuinucellate ovules have very small nucellus.
Ovules with subhypodermal sporogenous cell is called crassinucellate type. Normally these ovules have fairly large nucellus. Group of cells found at the base of the ovule between the chalaza and embryo sac is called hypostase and the thick-walled cells found above the micropylar end above the embryo sac is called epistase. The structure of ovule is given in Figure 1.7.
Types of Ovules
The ovules are classifid into six main types based on the orientation, form and position of the micropyle with respect to funicle and chalaza. Most important ovule types are orthotropous, anatropous, hemianatropous and campylotropous. The types of ovule is given in Figure 1.8.
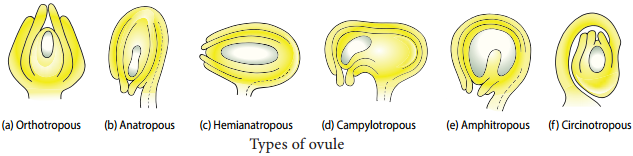
Orthotropous:
In this type of ovule, the micropyle is at the distal end and the micropyle, the funicle and the chalaza lie in one straight vertical line. Examples: Piperaceae, Polygonaceae.
Anatropous:
The body of the ovule becomes completely inverted so that the micropyle and funiculus come to lie very close to each other. This is the common type of ovules found in dicots and monocots.
Hemianatropous:
In this, the body of the ovule is placed transversely and at right angles to the funicle. Example: Primulaceae.
Campylotropous:
The body of the ovule at the micropylar end is curved and more or less bean shaped. The embryo sac is slightly curved. All the three, hilum, micropyle and chalaza are adjacent to one another, with the micropyle oriented towards the placenta. Example: Leguminosae In addition to the above main types there are two more types of ovules they are,
Amphitropous:
The distance between hilum and chalaza is less. The curvature of the ovule leads to horse-shoe shaped nucellus. Example: some Alismataceae.
Circinotropous:
Funiculus is very long and surrounds the ovule. Example: Cactaceae
Megasporogenesis
The process of development of a megaspore from a megaspore mother cell is called megasporogenesis.
As the ovule develops, a single hypodermal cell in the nucellus becomes enlarged and functions as archesporium. In some plants, the archesporial cell may directly function as megaspore mother cell. In others, it may undergo a transverse division to form outer primary parietal cell and inner primary sporogenous cell. The parietal cell may remain undivided or divide by few periclinal and anticlinal divisions
to embed the primary sporogenous cell deep into the nucellus.
The primary sporogenous cell functions as a megaspore mother cell. The megaspore mother cell (MMO) undergoes meiotic division to form four haploid megaspores. Based on the number of megaspores that develop into the Embryo sac, we have three basic types of development: monosporic, bisporic and
tetrasporic.
The megaspores are usually arranged in a linear tetrad. Of the four megaspores formed, usually the chalazal one is functional and other three megaspores degenerate. The functional megaspore forms the female gametophyte or embryo sac.
This type of development is called monosporic development (Example: Polygonum). Of the four megaspores formed if two are involved in Embryo sac formation the development is called bisporic (Example: Allium). If all the four megaspores are involved in Embryo sac formation the development is called tetrasporic (Example: Peperomia). An ovule generally has a single embryo sac. The development of monosporic embryo sac (Polygonum type) is given in Figure 1.9.
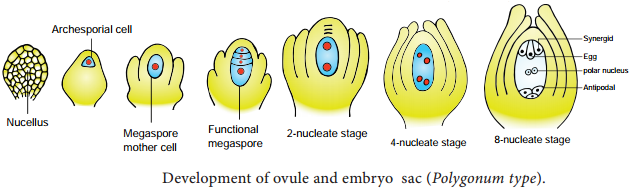
Development of Monosporic embryo sac.
To describe the stages in embryo sac development and organization the simplest monosporic type of development is given below. The functional megaspore is the first cell of the embryo sac or female gametophyte. The megaspore elongates along micropylar-chalazal axis.
The nucleus undergoes a mitotic division. Wall formation does not follow the nuclear division. A large central vacuole now appears between the two daughter nuclei. The vacuole expands and pushes the nuclei towards the opposite poles of the embryo sac.
Both the nuclei divide twice mitotically, forming four nuclei at each pole. At this stage all the eight nuclei are present in a common cytoplasm (free nuclear division). After the last nuclear division the cell undergoes appreciable elongation, assuming a sac-like appearance.
This is followed by cellular organization of the embryo sac. Of the four nuclei at the micropylar end of the embryo sac, three organize into an egg apparatus, the fourth one is lef free in the cytoplasm of the central cell as the upper polar nucleus.
Three nuclei of the chalazal end form three antipodal cells whereas the fourth one functions as the lower polar nucleus. Depending on the plant the 2 polar nuclei may remain free or may fuse to form a secondary nucleus (central cell).
The egg apparatus is made up of a central egg cell and two synergids, one on each side of the egg cell. Synergids secrete chemotropic substances that help to attract the pollen tube. The special cellular thickening called fiiform apparatus of synergids help in the absorption, conduction of nutrients from the nucellus to embryo sac. It also guides the pollen tube into the egg. Thus, a7 celled with 8 nuclei embryo sac is formed. The structure of embryo sac is given in Figure 1.10.
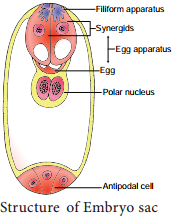
Pollination
Pollination is a wonderful mechanism which provides food, shelter etc., for the pollinating animals. Many plants are pollinated by a particular animal species and the flowers are modifid accordingly and thus there exists a co-evolution between plants and animals.
Let us imagine if pollination fails. Do you think there will be any seed and fruit formation? If not what happens to pollinating organisms and those that depend on these pollinating organism for the food? Here lies the signifiance of the process of pollination.
The pollen grains produced in the another will germinate only when they reach the stigma of the pistil. The reproductive organs, stamens and pistil of the flower are spatially separated, a mechanism which is essential for pollen grains to reach the stigma is needed. This process of transfer of pollen grains from the anther to a stigma of a flower is called pollination.
Pollination is a characteristic feature of spermatophyte (Gymnosperms and Angiosperms). Pollination in gymnosperms is said to be direct as the pollens are deposited directly on the exposed ovules, whereas in angiosperms it is said to be indirect, as the pollens are deposited on the stigma of the pistil.
In majority of angiosperms, the flower opens and exposes its mature anthers and stigma for pollination. Such flowers are called chasmogamous and the phenomenon is chasmogamy. In other plants, pollination occurs without opening and exposing their sex organs.
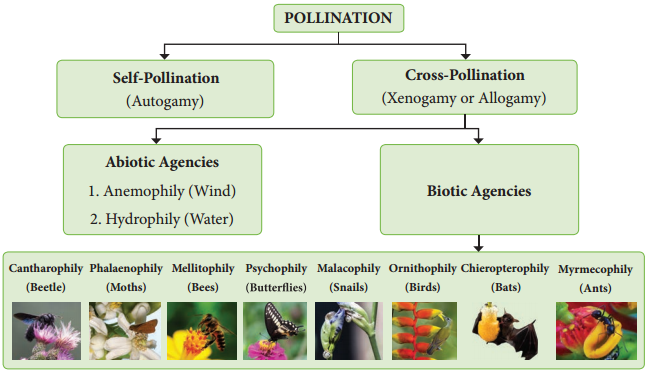
Such flowers are called cleistogamous and the phenomenon is cleistogamy. Based upon the flower on which the pollen of a flower reaches, the pollination is classified into two kinds, namely, self-pollination (Autogamy) and cross-pollination(Allogamy).

A. Self-pollination or Autogamy (Greek Auto = self, gamos = marriage):
According to a majority of Botanists, the transfer of pollen on the stigma of the same flower is called self-pollination or Autogamy. Self-pollination is possible only in those plants which bear bisexual flowers. In order to promote self-pollination the flowers of the plants have several adaptations or mechanisms. They are:
1. Cleistogamy:
In cleistogamy (Greek Kleisto = closed. Gamos = marriage) flowers never open and expose the reproductive organs and thus the pollination is carried out within the closed flower. Commelina, Viola, Oxalis are some examples for cleistogamous flowers. In Commelina benghalensis, two types of flowers are producedaerial and underground flowers.
The aerial flowers are brightly coloured, chasmogamous and insect pollinated. Th underground flowers are borne on the subterranean branches of the rhizome that are dull, cleistogamous and self pollinated and are not dependent on pollinators for pollination. (Figure 1.11).
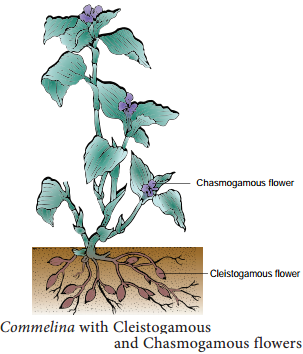
2. Homogamy:
When the stamens and stigma of a flower mature at the same time it is said to be homogamy. It favours self pollination to occur. Example: Mirabilis jalapa, Catharanthus roseus.
3. Incomplete dichogamy:
In dichogamous flowers the stamen and stigma of a flower mature at different time. Sometimes, the time of maturation of these essential organs overlap so that it becomes favourable for self-pollination.
B. Cross – pollination
It refers to the transfer of pollens on the stigma of another flower. The cross-pollination is of two types:
(i) Geitonogamy:
When the pollen deposits on another flower of the same individual plant, it is said to be geitonogamy. It usually occurs in plants which show monoecious condition. It is functionally cross-pollination but is generally similar to autogamy because the pollen comes from same plant.
ii. Xenogamy:
When the pollen (genetically different) deposits on another flower of a different plant of the same species, it is called as xenogamy.
Contrivances of cross-pollination
The flowers have several mechanisms that promote cross-pollination which are also called contrivances of cross-pollination or outbreeding devices. It includes the following.
1. Dicliny or Unisexuality
When the flowers are unisexual only crosspollination is possible. Thre are two types.
(i) Monoecious:
Male and female flowers on the same plant. Coconut, Bitter gourd. In plants like castor and maize, autogamy is prevented but geitonogamy takes place.
(ii) Dioecious:
Male and female flowers on different plants. Borassus, Carica and phoenix. Here both autogamy and geitonogamy are prevented.
2. Monocliny or Bisexuality
Flowers are bisexual and the special adaptation of the flowers prevents self-pollination.
(i) Dichogamy:
In bisexual flowers anthers and stigmas mature at different times, thus checking self-pollination. It is of two types.
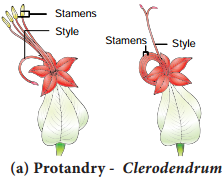
a. Protandry:
The stamens mature earlier than the stigmas of the flowers. Examples: Helianthus, Clerodendrum (Figure 1.12 a).
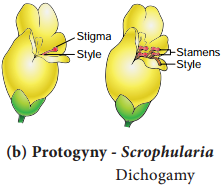
b. Protogyny:
The stigmas mature earlier than the stamens of the flower. Examples: Scrophularia nodosa and Aristolochia bracteata (Figure 1.12 b).
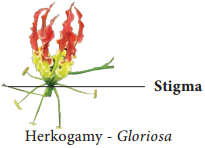
(ii) Herkogamy:
In bisexual flowers the essential organs, the stamens and stigmas, are arranged in such a way that self-pollination becomes impossible. For example in Gloriosa superba, the style is reflxed away from the stamens and in Hibiscus the stigmas project far above the stamens (Figure 1.13).
(iii) Heterostyly:
Some plants produce two or three different forms of flowers that are different in their length of stamens and style. Pollination will take place only between organs of the same length. (Figure 1.14)
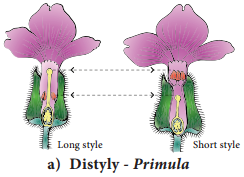
a. Distyly:
The plant produces two forms of flowers, Pin or long style, long stigmatic papillae, short stamens and small pollen grains; Thum-eyed or short style, small stigmatic papillae, long stamens and large pollen grains. Example: Primula (Figure 1.14 a).
The stigma of the Thum-eyed flowers and the another of the pin lie in same level to bring out pollination. Similarly the anther of Thum-eyed and stigma of pin ones is found in same height. This helps in effective pollination.
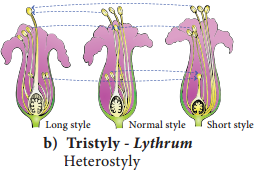
b. Tristyly:
The plant produces three kinds of flowers, with respect to the length of the style and stamens. Here,the pollen from flowers of one type can pollinate only the other two types but not their own type. Example: Lythrum (Figure 1.14 b).
(iv) Self sterility/Self- incompatibility:
In some plants, when the pollen grain of a flower reaches the stigma of the same, it is unable to germinate or prevented to germinate on its own stigma. Examples: Abutilon, Passiflra. It is a genetic mechanism.
Agents of pollination
Pollination is effcted by many agents like wind, water, insects etc. On the basis of the agents that bring about pollination, the mode of pollination is divided into abiotic and biotic. The latter type is used by majority of plants.
Abiotic agents
- Anemophily – pollination by Wind
- Hydrophily – pollination by Water
Biotic agents
Zoophily:
Zoophily refers to pollination through animals and pollination through insects is called Entomophily.
1. Anemophily:
Pollination by wind. The wind pollinated flowers are called anemophilous. The wind pollinated plants are generally situated in wind exposed regions. Anemophily is a chance event. Therefore, the pollen may not reach the target flower effctively and are wasted during the transit from one flower to another. The common examples of wind pollinated flwers are – grasses, sugarcane, bamboo, coconut, palm, maize etc.,
Anemophilous plants have the following characteristic features:
- The flowers are produced in pendulous, catkin-like or spike inflrescence.
- The axis of inflrescence elongates so that the flowers are brought well above the leaves.
- The perianth is absent or highly reduced.
- The flowers are small, inconspicuous, colourless, not scented, do not secrete nectar.
- The stamens are numerous, fiaments are long, exerted and versatile.
- Anthers produce enormous quantities of pollen grains compared to number of ovules available for pollination. They are minute, light and dry so that they can be carried to long distances by wind.
- In some plants anthers burst violently and release the pollen into the air. Example: Urtica.
- Stigmas are comparatively large, protruding, sometimes branched and feathery, adapted to catch the pollen grains. Generally single ovule is present.
- Plant produces flwers before the new leaves appear, so the pollen can be carried without hindrance of leaves.
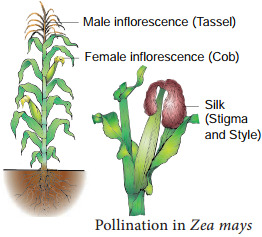
Pollination in Maize (Zea mays):
The maize is monoecious and unisexual. The male inflorescence (tassel) is borne terminally and female inflrescence (cob) laterally at lower levels. Maize pollens are large and heavy and cannot be carried by light breeze.
However, the mild wind shakes the male inflorescence to release the pollen which falls vertically below. The female inflorescence has long stigma (silk) measuring upto 23 cm in length, which projects beyond leaves. The pollens drop from the tassel is caught by the stigma (Figure 1.15).
Hydrophily:
Pollination by water is called hydrophily and the flowers pollinated by water are said to be hydrophilous (Example: Vallisneria, Hydrilla). Though there are a number of aquatic plants, only in few plants pollination takes place by water. The floral envelop of hydrophilous plants are reduced or absent. In water plants like Eichhornia and water lilly pollination takes place through wind or by insects.
There are two types of hydrophily, Epihydrophily and Hypohydrophily. In most of the hydrophilous flowers, the pollen grains possesses mucilage covering which protects them from wetting.
a. Epihydrophily:
Pollination occurs at the water level. Examples: Vallisneria spiralis, Elodea.
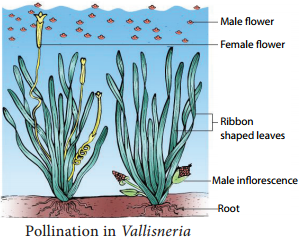
Pollination in Vallisneria spiralis:
It is a dioecious, submerged and rooted hydrophyte. The female plant bears solitary flowers which rise to the surface of water level using a long coiled stalk at the time of pollination. A small cup shaped depression is formed around the female flower on the surface of the water.
The male plant produces male flowers which get detached and flat on the surface of the water. As soon as a male flowers comes in contact with the female flower and pollination takes place, Stalk of the female flower coils and goes under water where fruits are produced. (Figure 1.16).
b. Hypohydrophily:
Pollination occurs inside the water. Examples: Zostera marina and Ceratophyllum.
Zoophily:
Pollination by the agency of animals is called zoophily and flowers are said to be zoophilous. Animals that bring about pollination may be birds, bats, snails and insects. Of these, insects are well adapted to bring pollination. Larger animals like primates (lemurs), arboreal rodents, reptiles (gecko lizard and garden lizard) have also been reported as pollinators.
A. Ornithophily:
Pollination by birds is called Ornithophily. Some common plants that are pollinated by birds are Erythrina, Bombax, Syzygium, Bignonia, Sterlitzia etc., Humming birds, sun birds, and honey eaters are some of the birds which regularly visit flowers and bring about pollination.
The ornithophilous flowers have the following characteristic features:
- The flowers are usually large in size.
- The flowers are tubular, cup shaped or urnshaped.
- The flowers are brightly coloured, red, scarlet, pink, orange, blue and yellow which attracts the birds.
- The flowers are scentless and produce nectar in large quantities. Pollen and nectar form the floral rewards for the birds visiting the flowers.
- The floral parts are tough and leathery to withstand the powerful impact of the visitors.
B. Cheiropterophily:
Pollination carried out by bats is called cheiropterophily. Some of the common cheiropterophilous plants are Kigelia africana, Adansonia digitata, etc.,
C. Malacophily:
Pollination by slugs and snails is called malacophily. Some plants of Araceae are pollinated by snails. Water snails crawling among Lemna pollinate them.
D. Entomophily:
Pollination by insects is called Entomophily. Pollination by ant is called myrmecophily. Insects that are well adapted to bring pollination are bees, moths, butterfles, fles, wasps and beetles. Of the insects, bees are the main flower visitors and dominant pollinators. Insects are chief pollinating agents and majority of angiosperms are adapted for insect pollination. It is the most common type of pollination.
The characteristic features of entomophilous flowers are as follows:
- Flowers are generally large or if small they are aggregated in dense inflrescence. Example: Asteraceae flowers.
- Flowers are brightly coloured. Th adjacent parts of the flwers may also be brightly coloured to attract insect. For example in Poinsettia and Bougainvillea the bracts become coloured.
- Flowers are scented and produce nectar.
- Flowers in which there is no secretion of nectar, the pollen is either consumed as food or used in building up of its hive by the honeybees. Pollen and nectar are the floral rewards for the visitors.
- Flowers pollinated by fles and beetles produce foul odour to attract pollinators.
- In some flwers juicy cells are present which are pierced and the contents are sucked by the insects.
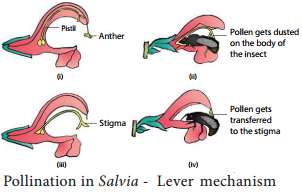
Pollination in Salvia (Lever mechanism):
The flower is protandrous and the corolla is bilabiate with 2 stamens. A lever mechanism helps in pollination. Each anther has an upper fertile lobe and lower sterile lobe which is separated by a long connective which helps the anthers to swing freely. When a bee visits a flower, it sits on the lower lip which acts as a platform. It enters the flwer to suck the nectar by pushing its head into the corolla.
During the entry of the bee into the flower the body strikes against the sterile end of the connective. This makes the fertile part of the stamen to descend and strike at the back of the bee. The pollen gets deposited on the back of the bee. When it visits another flower, the pollen gets rubbed against the stigma and completes the act of pollination in Salvia (Figure 1.17).
Some of the other interesting pollination mechanisms found in plants are a Trap mechanism (Aristolochia);Pit fall mechanism (Arum); Clip or translator mechanism.